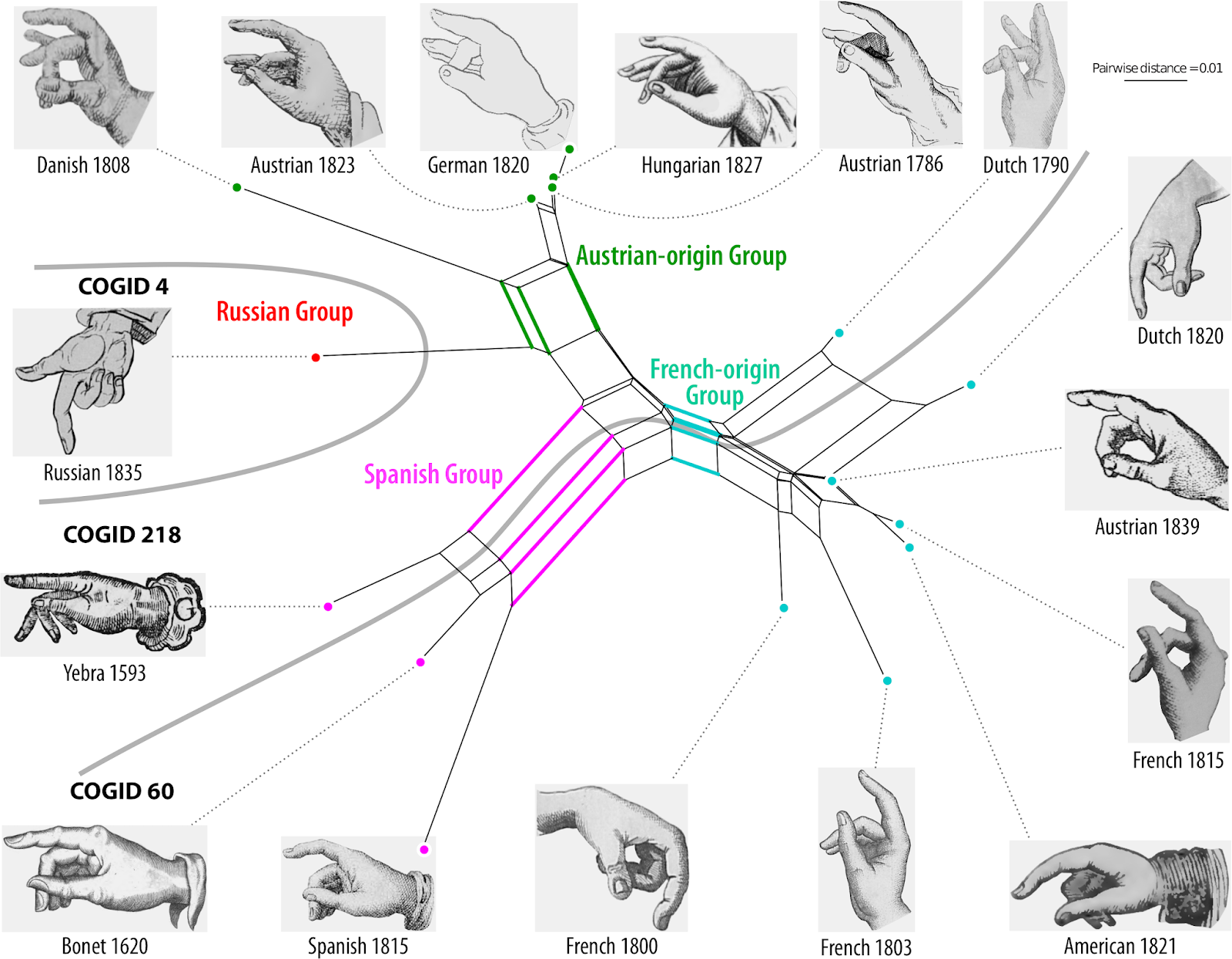
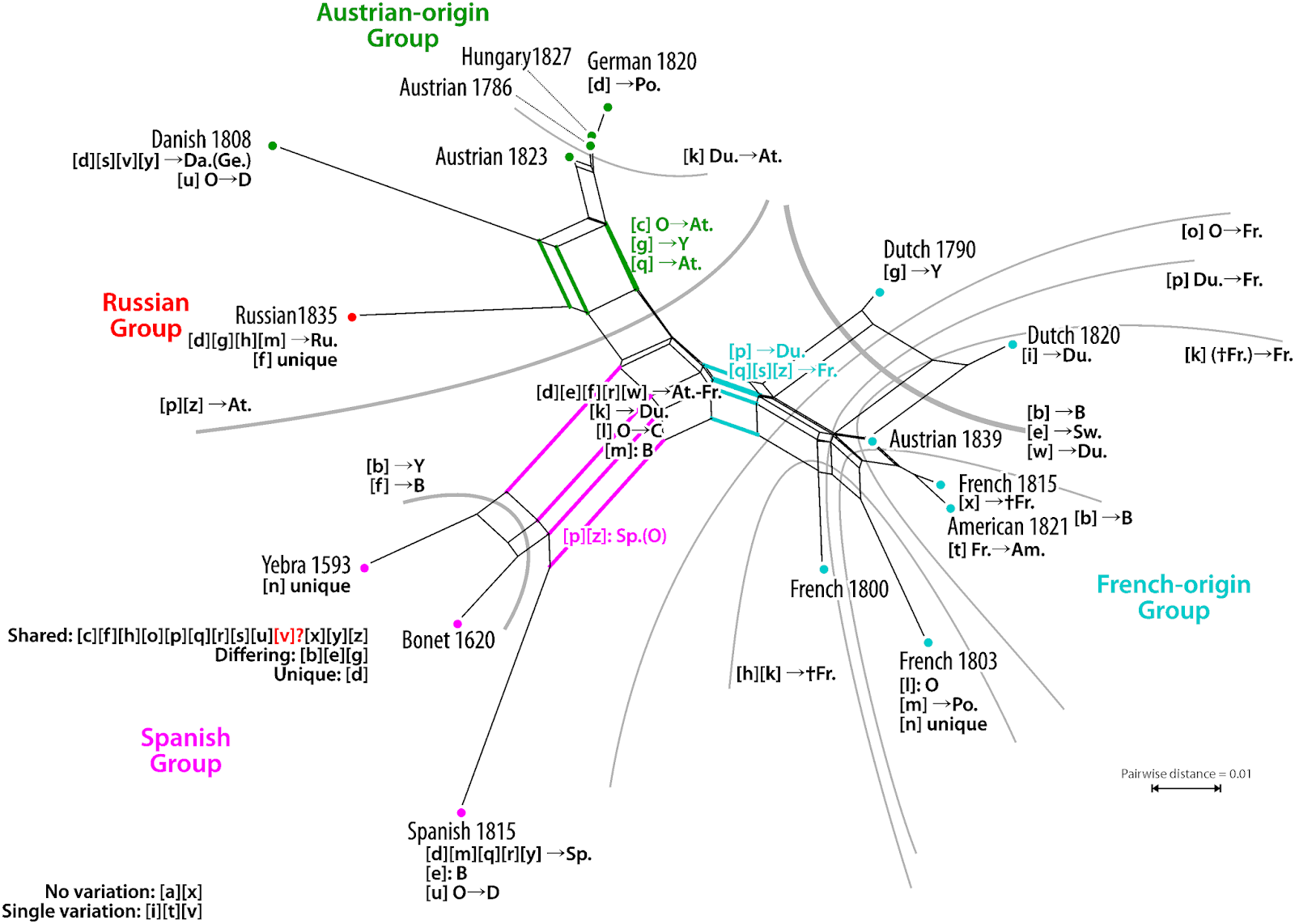
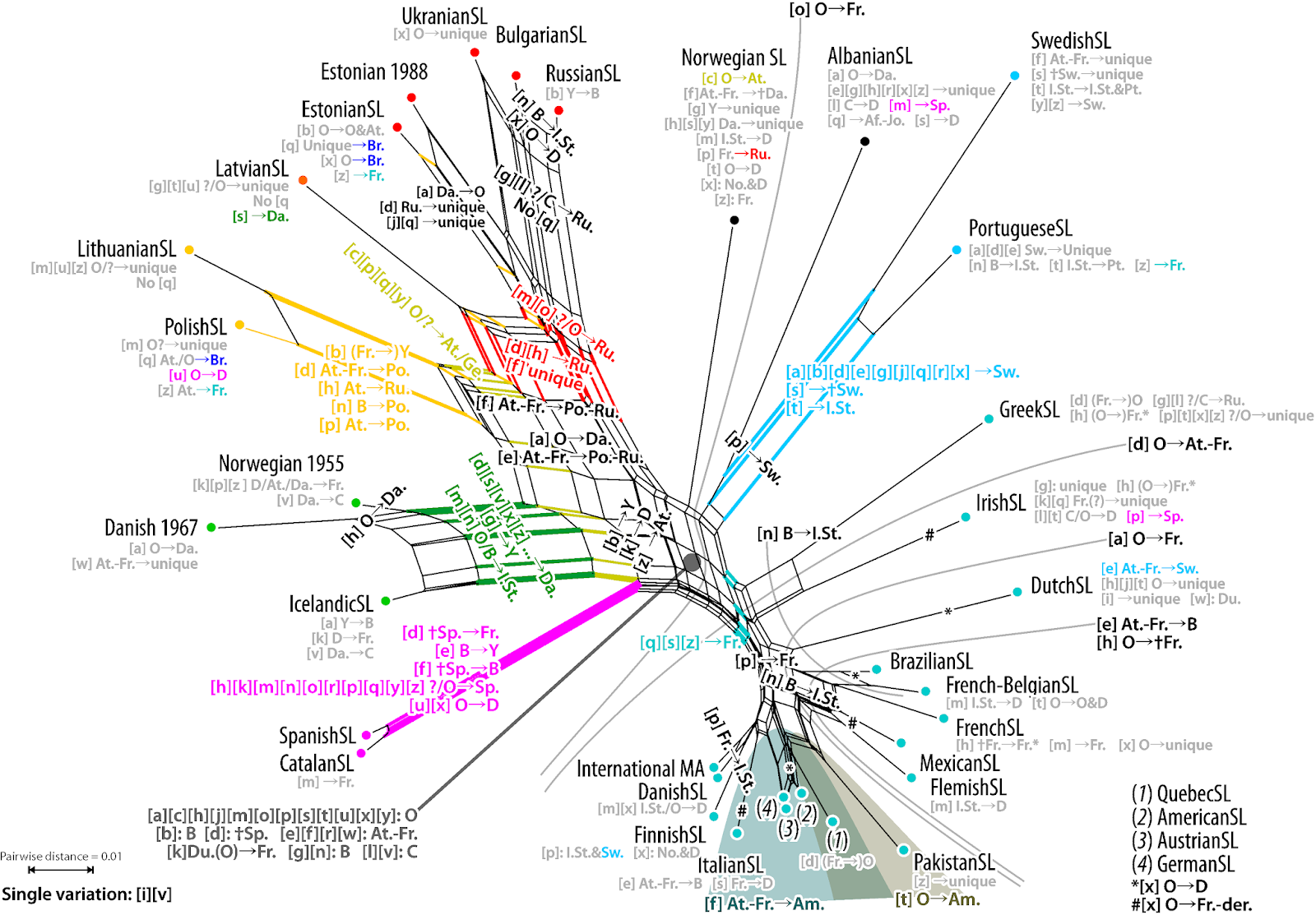
A nice example of this use is the Neighbor-net in a recent paper on Chinese oaks:
Yang J, Guo Y-F, Chen X-D, Zhang X, Ju M-M, Bai G-Q, Liu Z-L, Zhao G-F. Framework Phylogeny, Evolution and Complex Diversification of Chinese Oaks. Plants 2020: 1024.[Note: The paper is, from a purely methodological point-of-view, pretty well done, but has probably not experienced any real peer-review.**]
Oaks (Quercus L.) are ideal models to assess patterns of plant diversity. We integrated the sequence data of five chloroplast and two nuclear loci from 50 Chinese oaks to explore the phylogenetic framework, evolution and diversification patterns of the Chinese oak’s lineage. The framework phylogeny strongly supports two subgenera Quercus and Cerris comprising four infrageneric sections Quercus, Cerris, Ilex and Cyclobalanopsis for the Chinese oaks.None of this is new. My colleagues and I published an updated classification for oaks a few years ago (Denk et al. 2017) that took into account molecular phylogenies, and introduced the systematic concept referred to by Yang et al., and recently followed by a many-species global oak phylogenomic study (Hipp et al. 2020). All of this is based on nuclear data only, because any researcher who ever studies oak genetics soon realizes that the plastomes are largely decoupled from speciation processes, but are geographically highly constrained (eg. Simeone et al. 2016, Yan et al. 2019). This is the reason why oaks are indeed "ideal models to assess patterns of plant diversity" – they provide a worst-case scenario not the (trivial) best-case one.
As can be seen in the Yang et al. tree, members of section Ilex, a monophyletic lineage forming highly supported clades in trees based on nuclear data, are scattered all across the subgenus Cerris subtree. I have annotated a copy of this tree here.
 |
| Yang et al.'s fig. 1a, with some clades newly labeled for orientation |
Because of the plastid incongruence, the subgenus Cerris subtree has a wrong root (section Cylcobalanopsis diverged before sister sections Cerris and Ilex split). Also, the reciprocally monophyletic, genetically coherent sections Cerris (green) and Cyclobalanopsis (blue) are embedded in the much more diverse Ilex 3 and Ilex 4 clades. The remaining Ilex species are placed in two early diverged clades, which I have labeled Ilex 1 and Ilex 2 in the above tree (note: the taxon set only includes Chinese oak species). The only indication the tree gives that we have a data conflict issue is the low support (gray circles represent branches with Maximum likelihood bootstrap support > 60).
The network
When interpreting the phylogenetic implications of a Neighbor-net, we have to keep in mind that it is not a phylogenetic network in the strict sense (ie. displaying an evolutionary history), but is instead a meta-phylogenetic graph: a summary of incompatible splits patterns. Incompatibility can have different origins: reticulation, recombination, diffuse or poorly sorted signals, etc. Consequently, when looking at a Neighbor-nets and their neighborhoods (Splits and neighborhoods in splits graphs), we need to keep in mind what kind of data we used to calculate the underlying distance matrix in the first place.
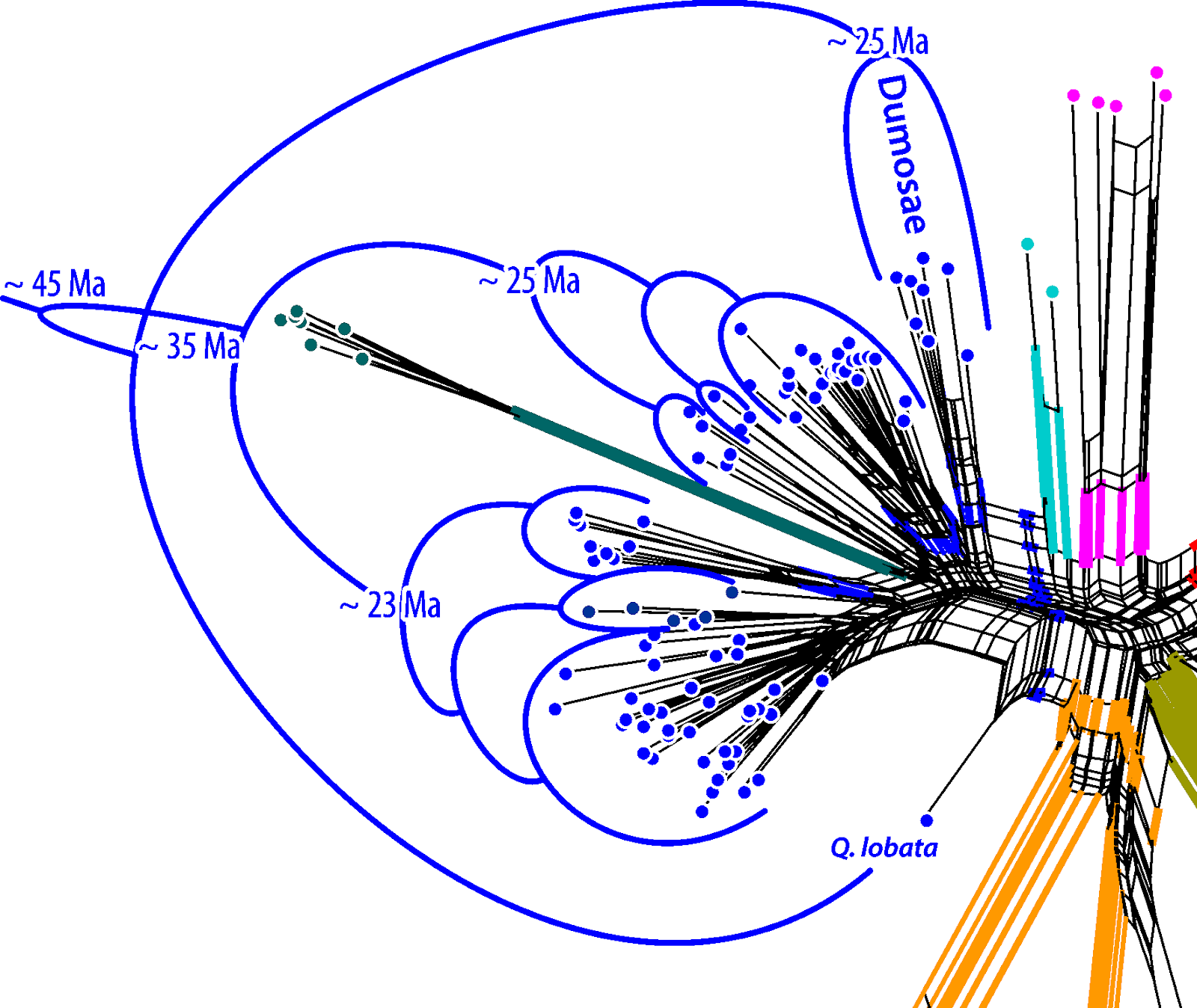
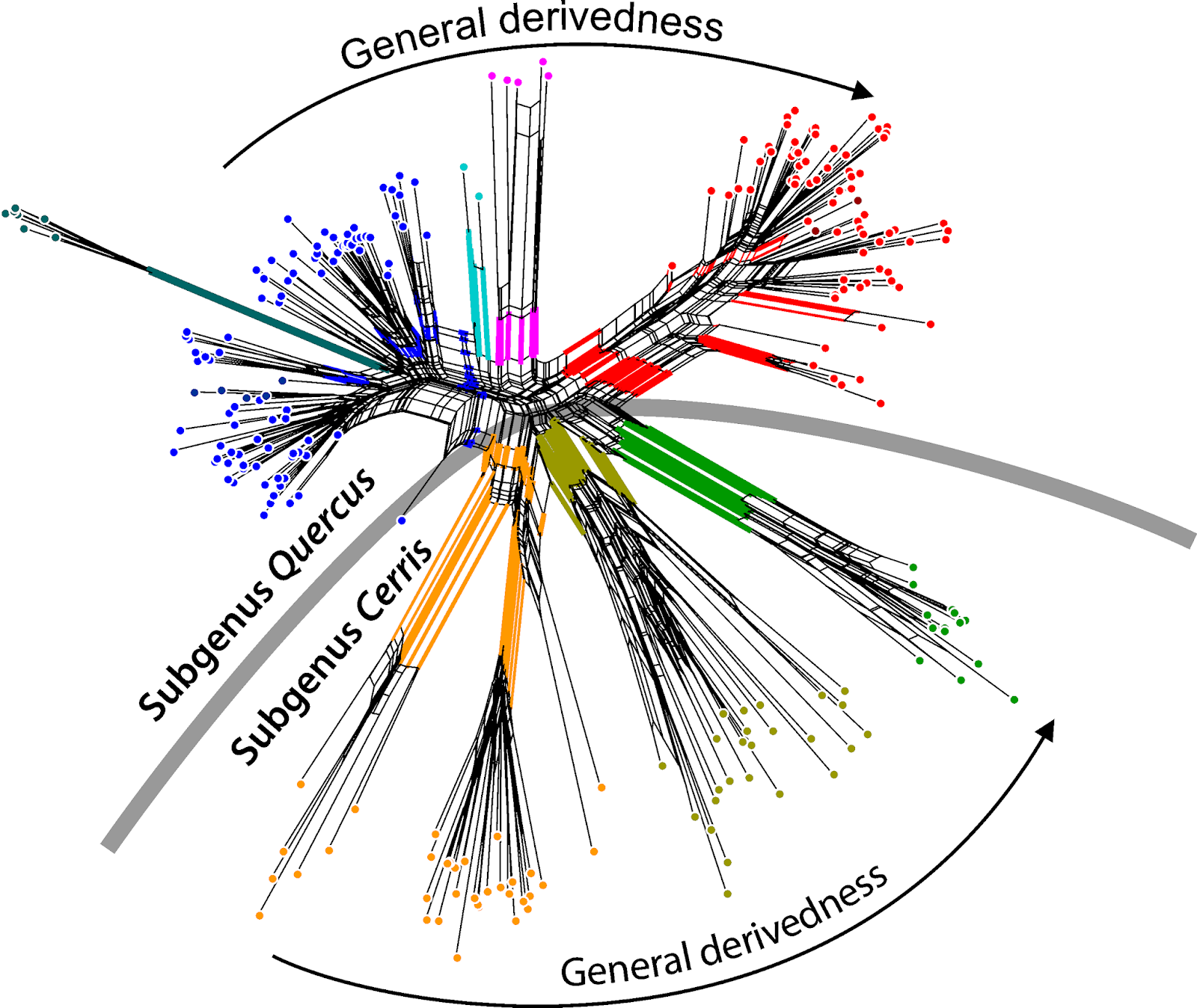
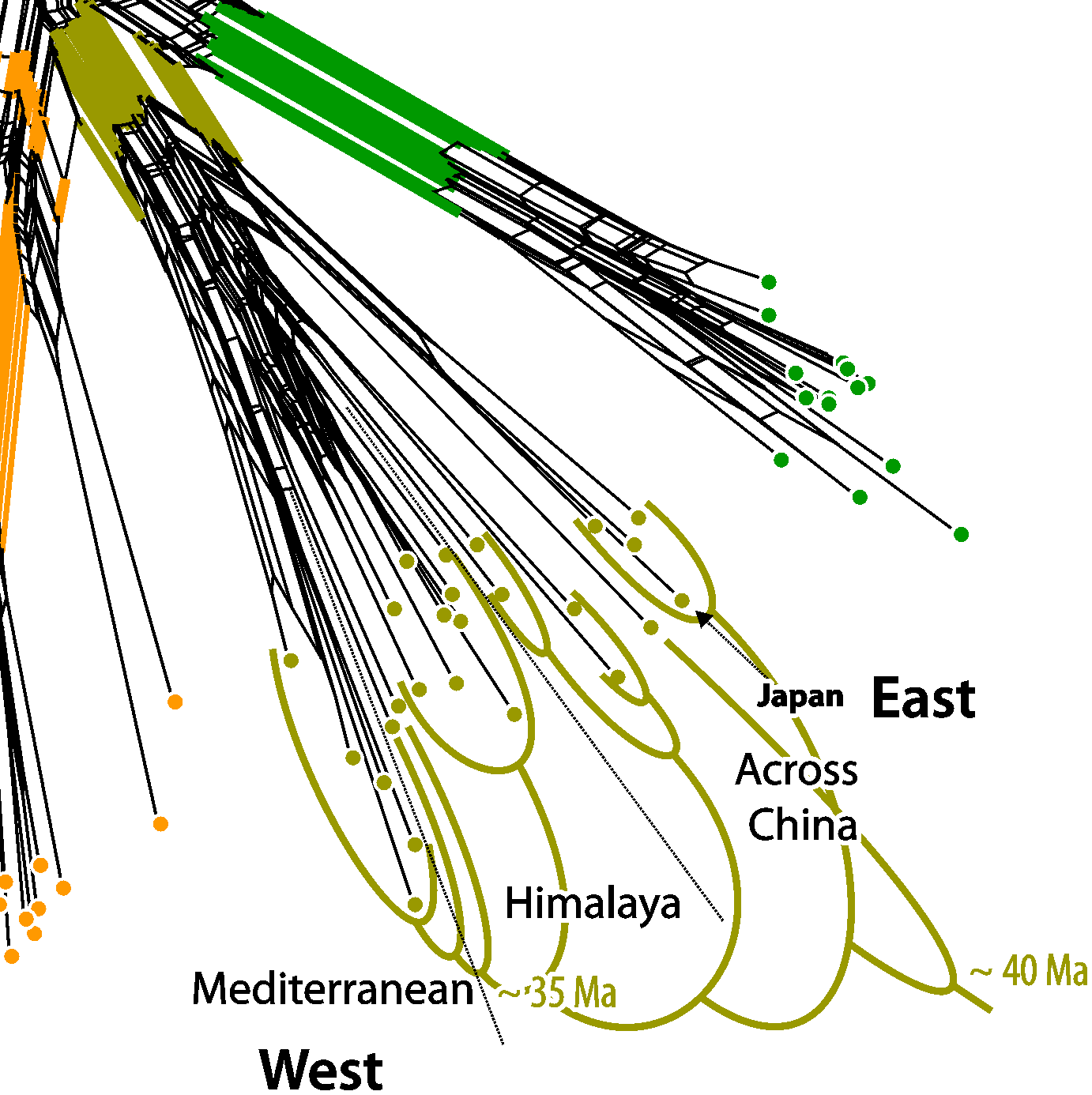
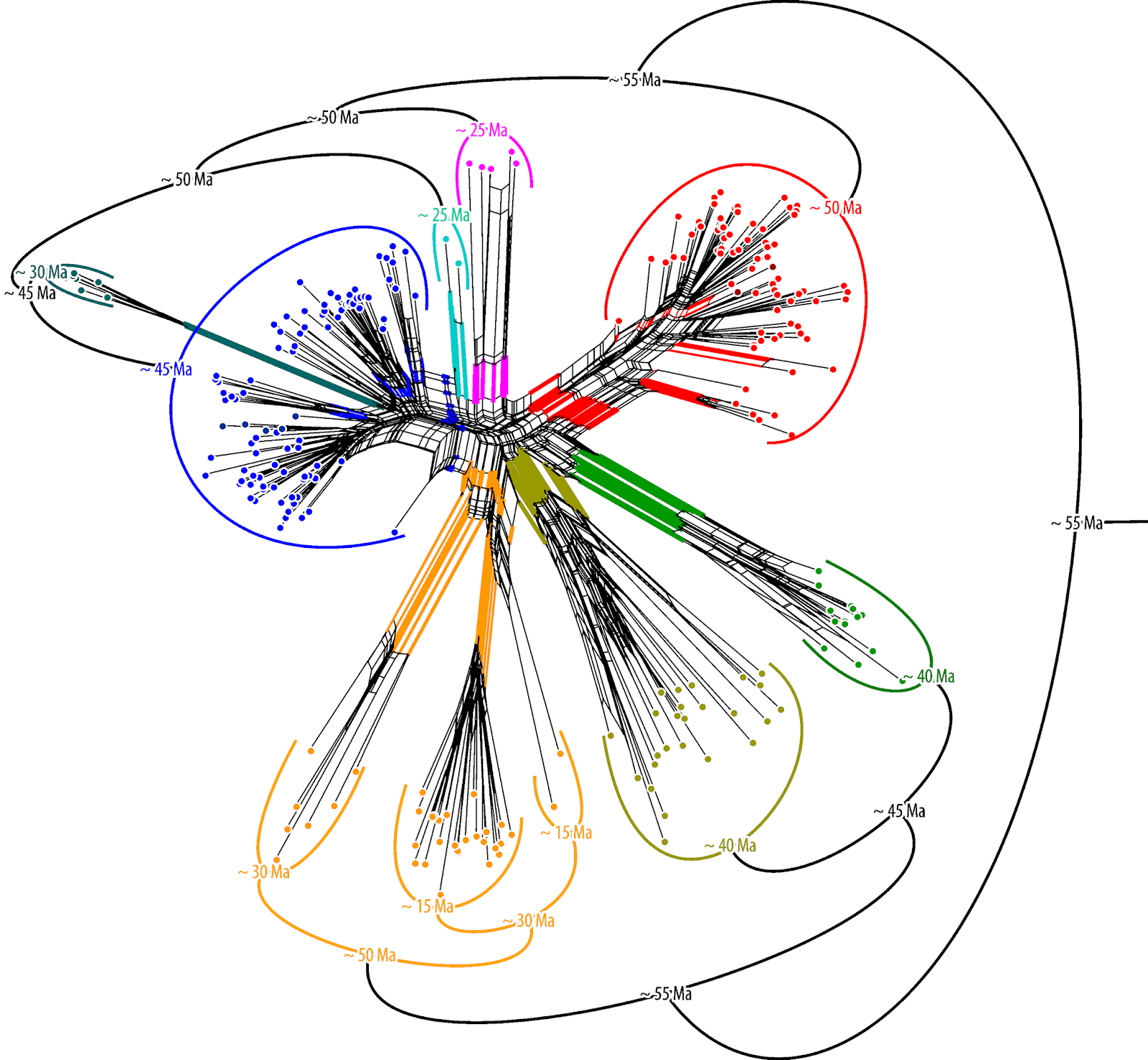
If the data follows two incongruent trees ("phylogenies"), as in this case for the oaks, the Neighbor-net has a good chance of capturing the incompatible splits of both genealogies. Here is the graph from the paper.
 |
| Wang et al.'s fig. 1b. |
The central inflated portion of the graph reflects the incongruence between the combined data sets: we have overlapping nuclear-informed and plastid-informed neighborhoods.
The authors' brackets (shown in black) refer to neighborhoods triggered by the two nuclear markers in the data set: these are neighborhoods reflecting the common origin and speciation within the oak lineages. We can even see that this signal, which is incompatible with all deep splits in the combined tree, is unambiguous in part of the data (the nuclear partitions): section Ilex spans out as a wide fan, but there is a relatively prominent edge bundle defining the according neighborhood (the blue split).
The net shows additional, even more prominent edge bundles defining partly overlapping or distinct neighborhoods (the red splits). These neighborhoods are represented as clades in Yang et al.'s phylogenetic tree (fig.1a). They write (p. 11 of 20):
However, the conflict between the two datasets seems to be recovered by the neighbor-net method in this study, as the neighbor-net network based on combined plastid–nuclear data strongly shows the presence of two subgenera and four infrageneric species groups for the Chinese oak’s lineage (Figure 1b).Interestingly, the authors nonetheless used the substantially incongruent combined data for downstream dating and trait mapping analysis (p. 7/20):
Bayesian evolutionary analyses provided a concordant infrageneric phylogeny for the Chinese oak’s lineage at the species level (Figure 2).This uses a taxon-filtered, obviously constrained (fixed) topology, fitted to the current synopsis outlined in Denk et al. (2017). [Note: the supplement includes the extremely incongruent nuclear and plastid trees, each of which has further incongruence issues because they combine fast- and very slow-evolving sequence regions.]
Postscript
More posts on oaks, plastid data and networks can be found here in the Genealogical World and in my Res.I.P. blog.
- Using median networks to understand evolution of genera (→ Example 3), Geneal. World Phyl. Networks, January 2018
- Reticulation at its best – an example from the oaks, Geneal. World Phyl. Networks, July 2018
- Comparing neighbour-nets and PCA graphs – the example of Mediterranean oaks, Geneal. World Phyl. Networks, March 2019
- Next-generation neighbor-nets, Geneal. World Phyl. Networks, April 2019
- Why you never should do a single-species plastid analysis of oaks, Res.I.P., September 2019
- When dating is futile – plastome-based chronograms for oaks, Res.I.P., October 2019
Cited papers
Denk T, Grimm GW, Manos PS, Deng M, Hipp AL. (2017) An updated infrageneric classification of the oaks: review of previous taxonomic schemes and synthesis of evolutionary patterns. In: Gil-Pelegrín E, Peguero-Pina JJ, and Sancho-Knapik D, eds. Oaks Physiological Ecology. Cham: Springer, pp. 13–38. Open access Pre-Print [major change: Ponticae and Virentes accepted as additional sections in final version].
Hipp AL, Manos PS, Hahn M, Avishai M, + 20 more authors. (2020) Genomic landscape of the global oak phylogeny. New Phytologist 229: 1198–1212. Open access.
Simeone MC, Grimm GW, Papini A, Vessella F, Cardoni S, Tordoni E, Piredda R, Franc A, Denk T. (2016) Plastome data reveal multiple geographic origins of Quercus Group Ilex. PeerJ 4:e1897. Open access.
Yan M, Liu R, Li Y, Hipp AL, Deng M, Xiong Y. (2019) Ancient events and climate adaptive capacity shaped distinct chloroplast genetic structure in the oak lineages. BMC Evolutionary Biology 19:202. Open access.
** The publisher, MDPI, thrives in the gray zone between predatory and accredited publishing. Originally included in the recently reactivated Beall's List (new homepage), it has been tentatively dropped (see the linked Wikipedia article; but see also this post by Mats Widgren). Personally, I have encountered articles published in MDPI journals only where the review process must have been, at least, strongly compromised. But it's always quick: Yang et al.'s paper was submitted July 24th, accepted August 12th, and published a day later. Three weeks is about the length of time that the editors of my first oak paper needed to find a peer reviewer at all.